2.6 Evidence from Molecular Biology and Genetics
2.6.1 DNA as Genetic Material. As was stated earlier (see I.1.4), genetic variability arising from mutation and recombination by natural selection serves as the raw material of evolution. Before the advent of molecular biology, very little was known concerning the nature and mode of action of the gene. Later, the identification of biological macromolecules in cells prompted many to explore the relationships between these substances and the Mendelian concepts (see I.1.3) of particulate genes. [145]
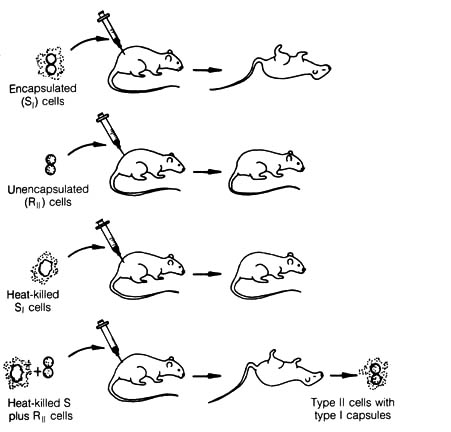
Figure 2.41. Bacterial Transformation. Transformation was discovered by F. Griffith, who noted that encapsulated (smooth or S) diplococci cause a fatal infection in mice, whereas nonencapsulated (rough or R) cells do not. Heat-killed S cells are likewise harmless, except when mixed with live R cells. In the latter case, a fatal infection can occur, and live cells having capsules characteristic of the S strain are found. Reprinted, with permission, from Dyson, R. D. Cell biology, a molecular approach. 1st ed. Boston: Allyn and Bacon, Inc.; 1974.
Figure 2.42. The Transforming Principle. When encapsulated diplococci were chemically fractionated, and the transformation experiment performed in vitro with each fraction one at a time, O. T. Avery and his colleagues found that DNA is the agent responsible for transformation. Reprinted, with permission, from Dyson, R. D. Cell biology, a molecular approach. 1st ed. Boston: Allyn and Bacon, Inc.; 1974.
The first experiment that paved the way for understanding the chemical
[146] nature of the gene was the discovery of transformation in
the bacteria Diplococcus pneumoniae by F. Griffith in 1928 (1).
The pathogenicity of the D. pneumoniae bacterium is associated with
the existence of a capsule. The capsule is a slimy material that
surrounds the cell and prevents phagocytosis by the white blood cells of
the host organisms. A nonpathogenic strain lacks the capsule and is easily
destroyed by host white blood cells. Griffith demonstrated that there were
stable materials from the heat-killed pathogenic strain of D. pneumoniae
(S
form) that could be transmitted and incorporated into the nonpathogenic
strain (R form). The materials could induce the latter to synthesize the
capsule and thereby transform the bacterium from a nonpathogenic to a pathogenic
strain (Figure 2.41). In 1944, Avery, Macleod, and McCarty (2)
identified the material responsible for this genetic transformation of
D.
pneumoniae
as DNA (Figure 2.42).
With the establishment of the chemical nature of the gene, the next important questions asked were (1) What is the chemical structure of DNA? and (2) How are genetic messages encoded in DNA? The answers to these two questions were provided by J. Watson and F. Crick (3, 4) and made a great impact on modern biology. The answers opened up the new discipline of molecular biology that presently is advancing rapidly and is influencing every area of modern biological thinking.
For a chemical model to account for the functions of a gene, it must
be able to show two characteristics: (1) the potential to duplicate itself
with exact fidelity so that each of the daughter molecules will be a replica
of itself and (2) the ability to carry coded information that specifies
a particular set of traits characteristic of a certain line of descent.
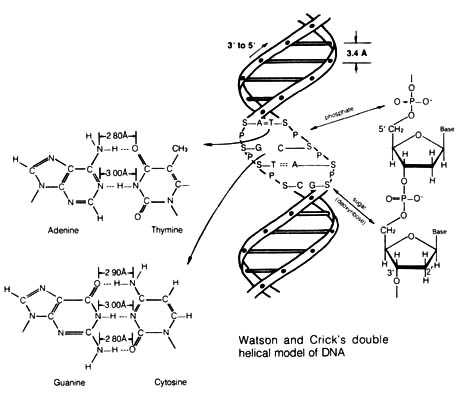
Watson and [147] Crick postulated a double helix model of DNA that satisfies
both of these criteria (Figure 2.43).
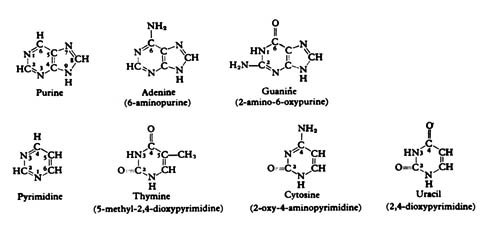
The structure of the double helix (DNA) can be viewed from several levels. The first and most important level is the bases, namely, the pyrimidines thymine and cytosine, and the purines guanine and adenine (Figure 2.44). They can form metastable paired configurations by hydrogen bonding (i.e., adenine paired with thymine via two hydrogen bonds; guanine paired with cytosine via three hydrogen bonds) (see Figure 2.43). The second level is the nucleoside. This is the combination of a single base with a 5 carbon (pentose) sugar having a 5 membered furanose ring. The absence of an oxygen at the 2' carbon position of the ring makes the sugar of the nucleoside deoxyribose. At the third level, there are nucleotides consisting of a nucleoside with an added phosphate group at the 5' position of the sugar. The fourth level involves the joining together of nucleotides by hooking up the 5' phosphate group of one with the 3' [148] position on the sugar ring of another, producing a long chain of nucleotides called a polynucleotides (Figure 2.45). The fifth and highest level involves two adjacent polynucleotide chains bound together by a long series of hydrogen bonds between the complementary bases forming a double helical structure.Figure 2.43. Base Pairing in DNA. A pyrimidine nucleotide (cytidine or thymidine) is always paired with a purine nucleotide (adenosine or guanosine), thus maintaining uniform overall dimensions in each pair. The hydrogen-bonding capabilities of the common form of the bases lead specifically to
and
pairing. Reprinted, with permission, from Dyson, R. D. Cell biology, a molecular approach. 1st ed. Boston: Allyn and Bacon, Inc.; 1974.
The double helical structure of DNA can readily account for replication. As soon as the double helix unwinds and separates, each strand serves as a template for the synthesis of the missing strand. Because of the specificity imposed on the pattern of hydrogen bonding between the adenine-thymine pair and the guanine-cytosine pair, DNA replication proceeds with high fidelity. During replication the parent strands of the DNA helix separate and a complementary daughter strand is synthesized on each parent strand. Thus two DNA molecules identical to the original molecule are produced (Figure 2.46).Figure 2.44. Purines and pyrimidines. Reprinted, with permission, from Dyson, R. D. Cell biology, a molecular approach. 1st ed. Boston: Allyn and Bacon, Inc.; 1974.
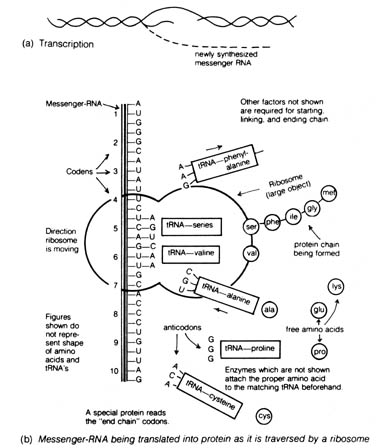
2.6.2 Gene Expression and the Genetic Code. The action of the genetic material in an organism can be summarized in the central dogma of molecular biology: DNA -> RNA -> protein. The information in the DNA is transcribed into the messenger RNA that is then translated into protein (Figure 2.47).
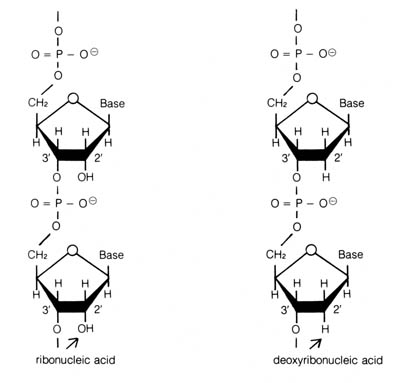
The RNA's or ribonucleic acid transcribed from the DNA is single stranded and contains the bases adenine, guanine, and cytosine as in DNA, but a new base uracil substitutes for thymine (Figure 2.44; 2.47). Also RNA has an oxygen at the 2' position of each sugar ring (see Figure 2.45) that is not present in DNA. Each different RNA is transcribed on only one of the DNA strands by complementary pairing facilitated by hydrogen bonding between bases (Figure 2.47a).
There are three types of RNA that can be transcribed from the DNA: messenger RNA, transfer RNA, and ribosomal RNA. Each transfer RNA carrying an amino acid makes contact with a messenger RNA at the ribosome [149] some through base pairing of a triplet base sequence (colon)
of the messenger RNA and a complimentary sequence (anticodon) of the transfer RNA (Figure 2.47b). Peptide bonds are formed between adjacent amino acids at the ribosomal level. By a process of translocation or the movement [151] of the ribosome along the messenger RNA, each successive codon of the messenger RNA binds with another transfer RNA that loses its amino acid to the growing amino acid polypeptide chain through peptide bond formation.
Figure 2.45. Presence of an oxygen in the 2' position of each sugar ring of the ribonucleic acid as contrasted with deoxyribonucleic acid.
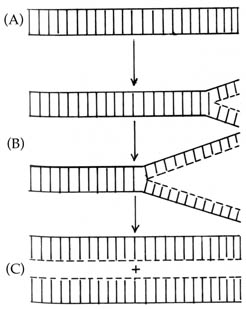
Figure 2.46. DNA replication. (A) Intact DNA double helix. The two horizontal lines represent the sugar phosphate backbone. The vertical lines represent hydrogen bonding between complimentary bases (see Figure 2.43). (B) DNA unwinds and separates. New DNA (---) is synthesized using the unwound strand of the parental DNA as a template. (C) Two daughter DNA double helices with one strand contributed by the parental molecule and one newly synthesized strand.
Figure 2.47. Transcription and translation. [150] As each colon or triplet of letters is read, a transfer-RNA molecule approaches which has an anticodon that will base-pair with those letters. This tRNA carries its matching amino acid which has been attached to it by its interpreter enzyme. As the tRNA is processed by the ribosome, the amino acid is joined onto the forming protein chain in the order called for by the mRNA sequence of code letters, which in turn was transcribed shortly before from the DNA master copy. This complex process takes place with fantastic speed and precision, and is remarkably similar in all living things known, from amebas to human beings. Recent evidence indicates that both transcription and replication may often be associated with cell membranes, including the endoplasmic reticulum. Adapted, with permission, from Coppedge, J. F. Evolution: possible or impossible? Grand Rapids, MI: Zondervan Publishing House; 1973.
By the elegant experiments of Nirenberg and Mattaei (5); Nishimura, Jones, and Khorana (6); and others, the genetic code was deciphered. The 20 amino acids present in proteins (Table 2.15) are specified by 64 triplet code words in DNA. These code words pass on the information via the codons in the messenger RNA and the anticodons in the transfer RNA, thereby specifying the amino acid sequence of the gene product. Since there are more code words than amino acids, a phenomenon known as degeneracy was observed, i.e., each amino acid is specified by more than one code word. Table 2.16 lists all the possible combinations of the triplet colons on the messenger RNA and the corresponding amino acids they specify.
There are also initiation and termination colons that are involved in the punctuation of the genetic message. The initiation colons GUG and AUG specify a particular amino acid named N-formylmethionine, which starts every polypeptide chain. However, when GUG and AUG are found in the middle of a polypeptide message, they specify instead valine and methionine respectively. The termination or nonsense colons are the UAA, UAG, and UGA colons that specify no amino acids. Thus the polypeptide chain falls off the ribosome as soon as it reaches one of these colons. Delimitated by the initiation and the termination codons, a gene is defined as the number of consecutive triplet codes on the DNA that determines the primary sequence of a polypeptide chain the so-called one-gene, one-polypeptide concept. This concept has been well documented in the genetic systems of bacteria and bacterial viruses and has provided modern biologists with the first operational tool to examine the interaction of genes.
The genetic code is probably universally applicable. The codons UUU, AAA, and CCC have been found to specify phenylalanine, lysine, and proline, respectively, in cell extracts prepared from a variety of different organisms ranging from bacteria to mammals (7). Purified messenger RNA encoding the protein hemoglobin extracted from rabbits was shown to be able to direct the synthesis of rabbit hemoglobin in frog oocytes, suggesting the colons in rabbits and frogs are translated the same way (8).
The apparent universality of the genetic code has been cited as evidence
that all living organisms arose from a single origin. After the early evolution
of the genetic code in organisms, it is thought to have remained constant
over a long period while other features of the organisms [152]
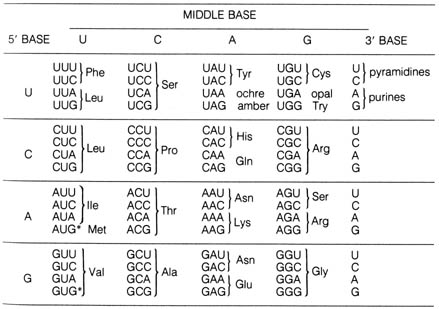
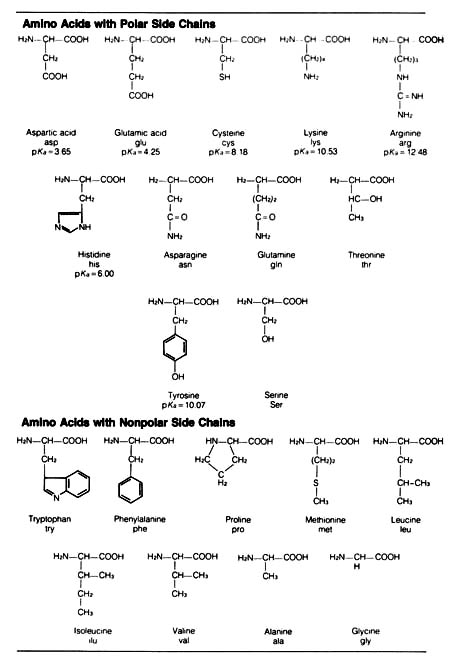
Table 2.15. The common amino acids.* 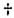
*The usual three-letter abbreviation is given, along with the pKa of side chain groups that may carry significant charge at physiological pH.
diverged [153] in subsequent evolution. However, the constancy of
the evolution of the genetic code may be challenged if examples of discrepancy
are discovered in future research. The presence of odd guanosine triphosphate
at the 5' end and polyadenosine monophosphate at the 3' end of the eucaryotic
messenger RNA with obscure functions may suggest that there are new mechanisms
of the expression of the genetic code yet to be discovered (9).
Alternatively, the universality of the code could be attributed to a Creator's
master design that enables all living organisms to operate under a similar
set of physiological conditions.
Proteins are ubiquitous components in the cell. They serve as the building blocks of cellular structure, and in the form of enzymes they also catalyze chemical reactions in the metabolic pathways of the cell. The functions of proteins are controlled by their structure that can be described at four levels: primary, secondary, tertiary, and quarternary.
Table 2.16. The genetic code.*![]()
*Note that in all cases but two (Tyr, Met), the third position may be occupied by either of the two purines or either of the two pyrimidines without changing the coding specificity. The terminator codonsUAA, UAG, and UGAstop amino acid incorporation and free the growing polypeptide chain. (The names ochre, amber, and opal refer to the mutant bacterial strains in which the action of these terminators were first studied.) Chain initiation begins with AUG or GUG, marked with an asterisk (*), either of which can code (in procaryotes) for N-formylmethionine in addition to the amino acid shown for it.
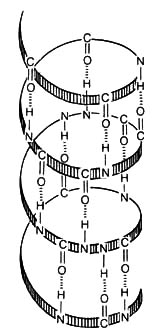
Figure 2.48. The alpha helix maintained by intrachain hydrogen bonding. [154]
The primary structure is the linear sequence of amino acids that are
joined together by peptide bonds to form a polypeptide chain. There are
20 naturally occurring amino acids in the living world 11 that are charged
ionic forms (those with polar side chains) and 9 that are neutral (those
with nonpolar side chains) under physiological conditions (Table 2.15).
Each species of protein has a unique amino acid sequence that essentially
determines its secondary, tertiary, and quarternary structure. One type
of secondary structure is the a helix configuration of a polypeptide chain
(Figure 2.48) (the envelope in Figure 2.49) formed by intrachain hydrogen
bonding. The tertiary structure is the apparent globular shape of the polypeptide
in solution when it coils together by intrachain, mostly noncovalent forces
between amino acid side chains. Proteins with more than one polypeptide
chain have quarternary structure, each chain being a subunit. The position
of each subunit is stabilized by noncovalent forces between chains (Figure
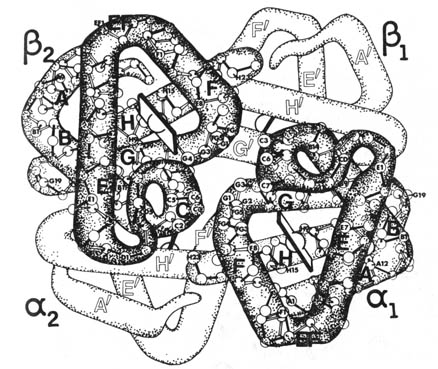
2.49). Hemoglobin (Figure 2.49), the oxygen-carrying protein in vertebrate
red blood cells, has a quarternary structure consisting of two ![]() and two
and two ![]() chains.
chains.
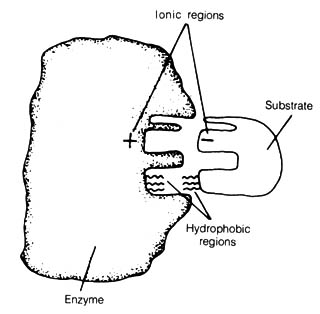
The function of any protein depends on the nature of its three-dimensional structure that is a reflection of the tertiary and quarternary [155] structure state. This can best be understood by the lock-and-key theory of enzyme function. The theory states that each enzyme (all enzymes are proteins) has a specific configuration that fits the substrate (Figure 2.50). If the functional three-dimensional structure of the enzyme is destroyed, the reaction catalyzed by the enzyme does not take place.Figure 2.49. Hemoglobin. Reproduced from Dickerson and Geis, slide set, Molecular structure of protein. By permission from Dr. Richard E. Dickerson, Department of Chemistry, California Institute of Technology, Pasadena.
Many inheritable phenotypic traits can be attributed to abnormal structure
of a single protein. The frequently cited example of a congenital disease
is sickle cell anemia (see I.3.2.l.c). This disease is caused by
the replacement of a single amino acid in the sixth position of the ![]() chain of hemoglobin. This replacement distorts the three-dimensional structure
of hemoglobin and causes the stacking of adjacent molecules in such a way
that the red blood cells carrying these distorted hemoglobins will become
sickle shaped and clump together in the absence of oxygen (Figure 2.51).
The sickle red blood cells last only half as long as normal cells, and
clumping of the sickle cells causes severe damage in vital organs. Therefore,
sickle cell anemic patients seldom live beyond 30 years of age. This [156]
has been called a molecular disease since the cause can be traced
to the distortion of a single molecule induced by a simple amino acid replacement.
chain of hemoglobin. This replacement distorts the three-dimensional structure
of hemoglobin and causes the stacking of adjacent molecules in such a way
that the red blood cells carrying these distorted hemoglobins will become
sickle shaped and clump together in the absence of oxygen (Figure 2.51).
The sickle red blood cells last only half as long as normal cells, and
clumping of the sickle cells causes severe damage in vital organs. Therefore,
sickle cell anemic patients seldom live beyond 30 years of age. This [156]
has been called a molecular disease since the cause can be traced
to the distortion of a single molecule induced by a simple amino acid replacement.
The production of enzymatic proteins is known to be a delicately regulated cellular process. The best-documented case of this type of gene expression is that of the operon model, which has been demonstrated in bacterial systems. An operon is a composite of several genes, usually clustering together in the bacterial chromosome and involved in similar functions. These genes are regulated by a single operator gene as the result of its interaction with the product of a regulatory gene. [157]Figure 2.50. Enzyme-Substrate Interaction. According to the lock-and-key theory, enzyme and substrate have complementary configurations. In this diagram, matching spatial conformations permit ionic and hydrophobic interactions to take place between the enzyme and its specific substrate. Reprinted, with permission, from Dyson, R. D. Cell Biology, a molecular approach. 1st ed. Boston: Allyn and Bacon, Inc.; 1974.
Figure 2.51. Photomicrograph of normal (dislike) and sickle (crescent-shaped) blood cells. Reprinted, with permission, from Lehninger, A. L. Biochemistry. 2nd ed. New York: Worth; 1975. Reprinted by permission from Walter Dawn, National Audubon Society, New York.
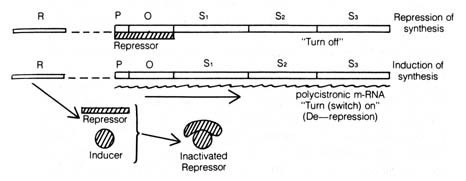
The best exemplified system of the operon is the lactose operon of the colon bacteria Escherichia coli (10) (Figure 2.52). The lactose operon is composed of three consecutive structural genes, each coding for a different enzyme:The OPERON Model OPERON: A single operator switches on a sequence of genes and it and the structural genes it controls comprise an integrated unit as both the physical and functional levels.Figure 2.52. The Jacob-Monod operon model for control of the synthesis of lactose-metabolizing enzymes. The repressor produced by the regulator gene (R) normally binds to the operator gene (O) and stops the transcription of the structural genes (S1, S2, S3) by blocking the RNA polymerase from binding to the promoter gene (P). In the presence of the inducer (lactose), the repressor is bound by it, and a conformational change takes place that inactivates the repressor from binding to the operator gene. Transcription of the structural genes gives rise to polycistronic messenger RNA (m-RNA) that is a continuous piece of RNA spanning across several genes. This m-RNA is then translated to form the three enzymes required before lactose can be used as an energy source.
The intriguing feature of this model is that the structural genes are expressed only when the operator gene is not bound by the repressor molecule. This can be brought about when an inducer, namely, lactose, is present and combines with the repressor to inactivate it. The enzymes encoded by the structural genes can thereby be synthesized and the inducer metabolized. As the lactose concentration decreases, more repressor [158] is freed to interact with the operator gene again and shut off the transcription of the structural genes. The repressor molecule has been isolated and identified. It is a so-called allosteric protein that can undergo changes in its three-dimensional structure when it is combined with small molecules such as lactose. The altered configuration of the repressor molecule accounts for the inability of the repressor to bind to the operator gene.
Thirty-one operons have been identified in the E. coli linkage map (11); therefore, the operon model is one of the very important modes of regulation in bacteria. Some of these operons may act in ways different from that of the lactose operon. However, all of them involve the action of regulator and operator genes on the structural genes. Whether the operon mechanism exists in eucaryotes is not known. Understanding the gene expression control mechanisms is difficult because the eucaryotic chromosome is a complex structure consisting of chromosomal proteins and chromosomal RNA and DNA. Some data, though limited, suggest that a type of operon exists (12). Mutation in regulator genes in the eucaryotic chromosomes are thought by some to be the raw material for molecular evolution. This concept will be considered further in the following section.
![]()
References 2.6
1. Griffith, F. J. Hyg. Camb. 27:113; 1928.
2. Avery, O. T.; MacLeod, C. M.; McCarty, M. J.
Exp. Med. 79:137; 1944.
3. Watson, J. D.; Crick, F. H. C. Cold Spring Harbor
Symp. Quant. Biol. 18:123; 1953.
4. Watson, J. D.; Crick, F. H. C. Nature. 171:464;
1953.
5. Nirenberg, M. W.; Mataei, J. H. Proc. Natl.
Acad. Sci. USA. 47:1588; 1961.
6. Nishimura, S.; Jones, D. S.; Khorana; H. G. J.
Mol. Biol. 13:302; 1965.
7. Watson, J. D. Molecular biology of the gene.
Menlo Park, CA: Benjamin; 1976: 374.
8. Lane, C. D.; Marbaix G.; Gurdon J. B. J. Mol.
Biol. 61:73; 1971.
9. Watson, J. D. Molecular biology of the gene.
482-83.
10. Jacob, F.; Monod, J. J. Mol. Biol. 2:318;
1961.