2.3 Evidence from Physical Anthropology (continued)
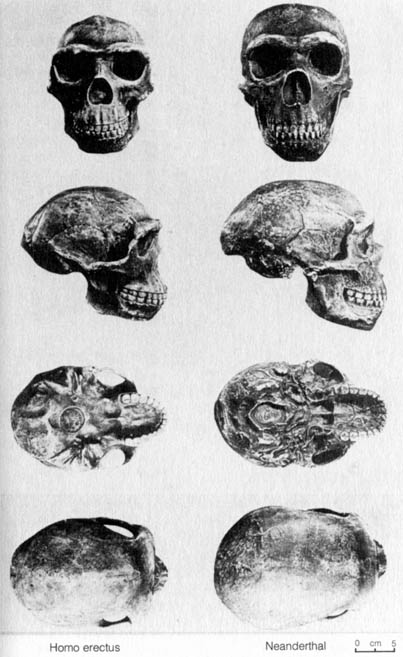
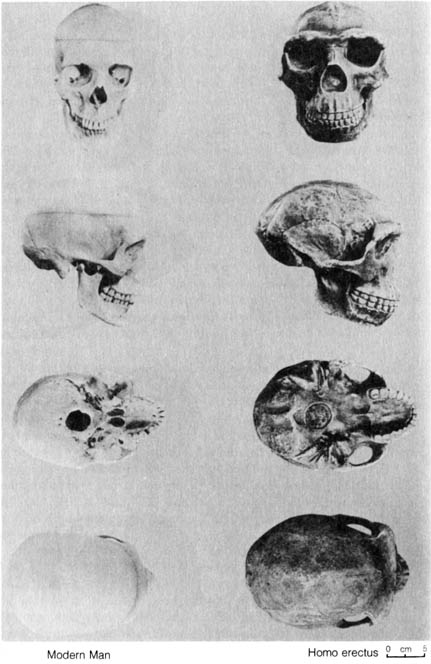
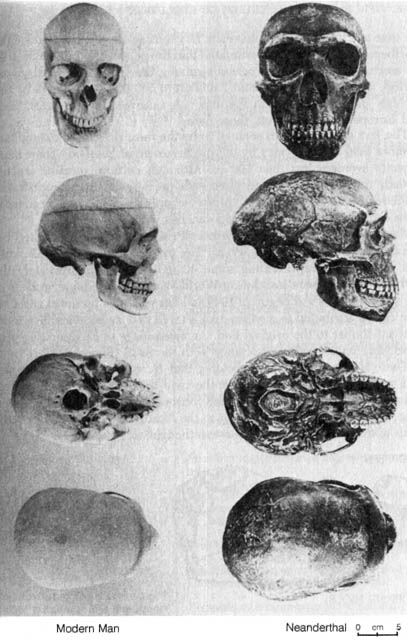
2.3.4 Homo erectus and Homo neanderthalensis. H. erectus and H. neanderthalensis are the two better documented early human fossils that trace the history of human beings back to the glacial periods. In addition to the large cranial capacities of these two species, the cultural remains also lend much support to their being human fossils. H. erectus was found not only with tools made of chipped stone and bone but also charred deer bone, a fact that strongly suggests they had learned to cook with fire. H. neanderthalensis had better-quality stone tools and buried their dead. Figures 2.20, 2.21 and 2.22 compare H. erectus with H. neanderthalensis, H. erectus with modern humans, and H. neanderthalensis with modern humans, respectively.
a) Homo erectus Discussed. The original find of H. erectus
was
made in Java in 1890-91 and includes fragments of a skull and a femur.
It was designated as Pithecanthropus erectus. Subsequently, various
skulls and jaws were found in Java, China, Hungary, Germany, Algeria, Morocco,
South Africa, and Tanzania (Table 2.12). The best documented among them
was the Sinanthropus pekinensis with many fragments of 15 skulls
and 11 mandibles found near Peking, China. It was renamed H. erectus
pekinensis and dated by an amino acid racemization technique (23)
to be around 300,000 to 500,000 years old. Analyzed fossil fauna associated
with the human fossils are suggestive of the climatic change of [109]
[112] the time and have helped to correlate H. erectus with the glaciation of the Northern Hemisphere. It is estimated that the period of time occupied by H. erectus appears to be a period spanning the first and second glacial period, and this is roughly from 1 million to 500,000 years ago (24). The absolute dating estimates on the materials uncovered from Africa, Java, and Europe also corroborate these dates.Figure 2.20. Comparison of four views of Homo erectus with a Neanderthal skull cast. Reprinted, with permission, from Kelso, A. J. Physical anthropology. Philadelphia: J. B. Lippincott & Co.; 1970. [110]
Figure 2.21. Comparison of four views of a modern human skull with a Homo erectus. Reprinted, with permission, from Kelso, A. J. Physical anthropology. Philadelphia: J. B. Lippincott & Co.; 1970. [111]
Figure 2.22. Comparison of four views of a modern human skull with a Neanderthal skull cast. Reprinted, with permission, from Kelso, A. J. Physical anthropology. Philadelphia: J. B. Lippincott & Co.; 1970.
The Java group of fossils seemed to be the most primitive, dating back to more than 710,000 years by the potassium-argon method. Their brain capacity averages less than 900 cc. Although cultural remains are not directly found in the original place with the Java fossils, circumstantial evidence indicates the oldest chopper-chopping stone tools recovered from central Java may be contemporaneous with the fossils.
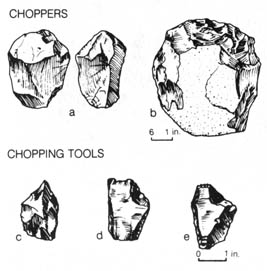
The H. erectus found in a cave near Peking was the best studied because of the abundance of materials. The original find included 15 skulls and other bones representing some 40 individuals. Unfortunately the early discoveries were lost during World War II. Subsequent work from the same deposits was begun in 1949 and has yielded a humerus and tibia that can be attributed to modern man, as well as a jawbone and five teeth that are similar to the original find. An assemblage of chopper-chopping tools were uncovered in association with the fossil (Figure 2.23).
There is strong evidence indicating that H. erectus knew how
to use and control fire. The evidence at the cave deposits includes fireplaces
found at several locations and many burned or charred fossil bones of other
animals presumably brought to the location by H. erectus. On the
basis of this evidence, H. erectus was thought to be a hunter.
Figure 2.23. Comparison of choppers with chopping tools. Reprinted, with permission, from Kelso, A. J. Physical anthropology. Philadelphia: J. B. Lippincott & Co.; 1970. [113]
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
NOTE: Adapted, with permission, from Kelso, A. J. Physical anthropology. Philadelphia: J. B. Lippincott & Co.; 1970.
The evidence of H. erectus in Europe is meager. No artifacts have been recovered in the sites where fossils were located. However, the associated [114] fauna indicate that these beings may have lived during the first interglacial period.
Artifacts discovered with the African H. erectus are indicative of the advanced lower Paleolithic tradition. The Olduvai find in Tanzania is associated with early hand ax archaeological remains. The H. erectus fossil found near the bottom of Bed II is dated to one million years ago.
A recently unearthed cranium of H. erectus from Lake Ndutu in northern Tanzania indicates a link between H. erectus and H. sapiens. Both the cranial features of H. erectus and H. sapiens are represented in this skull. It was dated at 500,000 to 600,000 years by the amino acid racemization method. It is probably the transitional form between H. erectus and H. sapiens (25, 26).
Collectively, H. erectus can be described as of moderate but erect stature, as shown by the straight limb bones, broad hip bones, and the position of the occipital condyle. The relative proportions of arms and legs are like those of modern humans. The forehead is retreating, and the jaw is projecting, though both to a much lesser extent than in the ape. The dentition is essentially that of modern humans. H. erectus was distributed throughout the Old World from about 1 million to 500,000 years ago, although not without exception (i.e., it has been claimed that skulls buried merely 10,000 years ago in Australia showed typical H. erectus characteristics) (27). They made the chopper-chopping type or the hand-ax type of tool, both presumably having the same subsistence function. Their appearance in the fossil record seems to be associated with a shift of mammalian fauna toward a warmer and moister climate. H. erectus was essentially a hunter and cave dweller, and at least at one site found to date it appears he used fire for food preparation.
b) Homo Neanderthalensis Discussed. The first Neanderthal fossil was discovered in 1848 at Gibraltar but was not thought to be significant in the scientific community. Eight years later, a similar skull together with a few ribs and limb bones were found in a cave in the Neanderthal Valley of Germany. They were the first bones to draw the attention of anthropologists. Since that time a large collection of similar fossils, some of them quite complete; have been uncovered in the Republic of South Africa, Zambia, France, Belgium, Italy, Yugoslavia, Israel, Uzbekistan, and Java (Table 2.13).
From the skeletal remains a rather complete picture of Neanderthal beings can be constructed. Their skull was thick-boned, with prominent eyebrow ridges and receding forehead. The roof of their skull was flat. However, their average cranial capacity of 1450 cc exceeds that for some modern humans (Figure 2.22). [115]
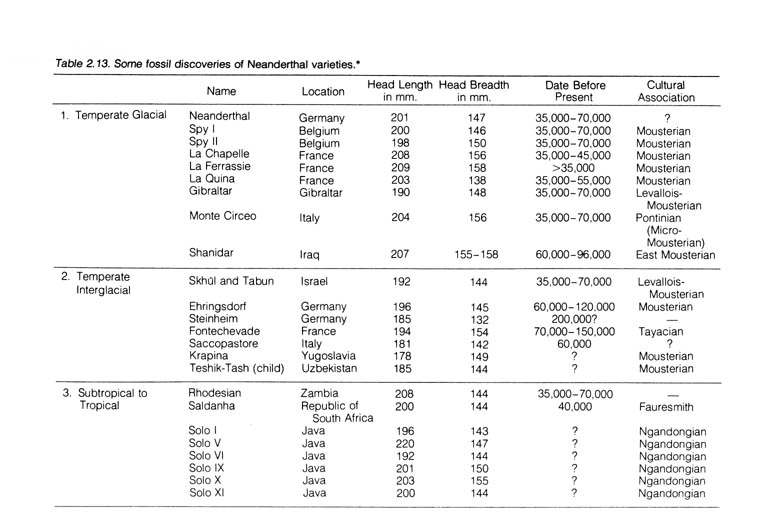
*NOTE: Reprinted, with permission, from Kelso, A. J. Physical anthropology. Philadelphia: J. B. Lippincott & Co.; 1970.
[116] The quality of the stone tools uncovered with the fossils
is indicative of middle to upper Paleolithic culture. Since they were scattered
through a period from approximately 150,000 years to 30,000 years ago,
they can be categorized into three groups according to the climates where
they lived: interglacial, glacial, and tropical. It can be safely concluded
that the glacial group of Neanderthal beings were hunters, whereas it seems
probable that the interglacial and the tropical groups were more vegetarians
than hunters. The interglacial groups were known to practice burials and
to place implements in the graves. The burials may have been in a single
grave, but multiple burials resembling a family cemetery were also uncovered.
A flint tool kit is frequently found buried with a dead male. One young
Neanderthal child was found buried with the horns of four ibexes, the great
mountain goat. The most impressive burial site was uncovered from the Shanidar
cave in the Zagros Mountains in northern Iraq. The skull of the burial
seemed to belong to a male around 30 years old who was killed by some strong
blows that badly crushed his head. He was buried on a bed with a blanket
made up of flowers, as evidenced by pollen analysis. This elaborate burial
rite leaves the impression that H. neanderthalensis believed in
life after death. It is also amazing to note that the man buried showed
evidence of a withered right arm that had been amputated, reattached, and
then healed.
Reconstruction from early finds depicted Neanderthal beings as persons with a stooped posture. The discoveries of more Neanderthal remains changed the picture. It is now apparent that the old conception of a Neanderthal being had been based on a misinterpretation of the study of an arthritic skeleton. Closer examination of the remains of other Neanderthal finds suggests that H. neanderthalensis was not much taller than five feet but stood with an upright posture.
Because of the advanced skeletal features of Neanderthal beings and their advanced stone tools and burial rites, H. neanderthalensis has been treated by many as a subspecies of modern humans Homo sapiens neanderthalensis. This interpretation was substantiated by the discovery in caves of Mount Carmel in Israel of a possible intermediate between the two species.
Since modern humans (Homo sapiens) appear abruptly at around 40,000 years ago at the advent of Upper Paleolithic culture when the Neanderthal beings disappeared from Europe, it has been speculated that H. sapiens neanderthalensis is the direct ancestor of modern humans. Except for the recent find of a possible intermediate between H. erectus and H. Sapiens (21, 22), there is no evidence against this interpretation. However, there are still unresolved questions raised in several finds (e.g., [117] the Swanscombe skull and the Steinheim and Ehringsdorf skulls) concerning the antiquity of H. sapiens. Therefore, no conclusive human lineage can be established to date.
However, it seems reasonable to assume that the earliest fossil that can be placed in direct human lineage, according to the present evidence, is H. erectus, who lived around 1 million to 500,000 years ago, and possibly the KNM-ER-1470 skull. But the identity of this latter skull has to await excavation of similar remains and, more importantly, the excavation of cultural materials associated with this being so that the cultural capacity of the fossil can be evaluated.
In summary, Ramapithecus and Australopithecus are thought by many to be the ancestral stock of H. erectus. H. erectus is considered by many to be the forerunner of H. Sapiens (modern humans). However, the presence of conflicting interpretations of the fossil material and the scarcity of cultural artifacts does not permit conclusive statements to be made about the evolutionary position of these forms.
![]()
References 2.3